
 m.quanpro.cn
m.quanpro.cn
媒体:生态学习资料 作者:W. Marchan
专业号:孙莹
2024/3/8 9:44:43
来自欧洲温带原始森林的挪威云杉和欧洲山毛榉树苗的加速生长速度与气候变暖有关
01
—
文章信息
第一作者:W. Marchand
通讯作者:W. Marchand
作者单位:捷克布拉格生命科学大学林业和木材科学学院森林生态系
02
—
摘要
森林的全球变化结果将受到幼树人口统计学的强烈影响。我们使用了来自欧洲广泛的森林清查图网络的数据,以量化气候因素与两种生态优势物种(挪威云杉和欧洲山毛榉)幼树生长率之间的关系。我们用原始森林中约17500棵树木的年径向生长测量值拟合非线性回归模型,以研究个体对温度和供水措施的敏感性。我们控制了多种潜在的混杂因素,包括个体发育、资源竞争以及人为氮和硫的沉积。与山毛榉相比,云杉的生长潜力显著提高,反映了物种与环境驱动因素的特定关系。水分可用性的下降更强烈地限制了云杉的生产力,而山毛榉对观察到的水分限制水平具有明显的耐受性。变暖促进了这两个物种的生长,但超过热最佳条件的生长季节温度限制了木材生产。我们发现,自19世纪初以来,幼树的重建年增长率有长期的积极趋势,这可能是由工业时代的变暖所驱动的。然而,我们的研究结果表明,由于热阈值和对水胁迫的不同耐受性,持续变暖和未来更普遍的干旱可能最终抑制生长。因此,全球变化因素可能会影响欧洲未来的物种丰度模式、生物量生产和森林碳汇能力。
03
—
引言
自然生态系统目前正经历着前所未有的环境条件变化,这与人为驱动的气候变暖有关。更具体地说,有记录的年平均气温上升预计将持续,并可能在不久的将来加速。位于高纬度和高海拔地区的森林可能特别容易受到温度驱动的影响,包括结构和功能的变化,因为这些地区的变暖趋势相对于全球平均水平更为陡峭。例如,在东欧喀尔巴阡山脉,那里的树木生长历来受到低温和生长季节短的限制,热等温线在海拔上逐渐向上移动。因此,生长季节长度正在增加,从而可能提高森林生产力。然而,气候变化对整个生态系统净生产力的影响仍然不确定。高海拔地区生产力的提高可能会被低海拔地区的相应下降所抵消,在低海拔地区,气候变暖可能超过某些树种的生态最佳值。次优气候条件也可能导致局部物种灭绝或物种向上迁移,以匹配合适栖息地的减少。我们对森林生产力随之变化的理解因先前记录的春季酚学进展可能越来越受到冬季低温不足的限制而进一步模糊,这影响了植被发育的冷藏需求。全球变化因素对森林影响的持续不确定性突出表明,需要更有力地量化环境变化与树木人口学之间的关联。
木本物种每年形成的生长增量被广泛用于研究树木对环境变化的反应。这些自然档案对于评估植物对气候条件年际变化的敏感性特别有用。例如,树木个体大小的年增长在一定程度上取决于外部条件与物种生态需求的匹配程度。在温度有限的环境中,气候变暖导致的年度碳同化增加预计将转化为更广泛的生长环。或者,更频繁的干旱和热浪,以及晚春霜冻的增加,可能会抑制树木生长,并抵消与变暖相关的春季物候学进展的任何有益影响。
树种具有不同的特征,这些特征会对环境约束产生独特的反应。例如,相对于欧洲山毛榉(Fagus sylvatica L.),挪威云杉(Picea abies(L.)H.Karst)的气孔调节策略更为保守。因此,水分胁迫最初可能会在更大程度上减少云杉的碳同化,但也会保护该物种导水木质部细胞的完整性和功能。在山毛榉等阔叶树种中,每年春天都会更换整个光合装置,这使得极端温度或缺水对最近形成的冠层叶片造成的任何损害都对树木生长极为不利。山毛榉的叶片潮红也发生在生长季节相对较早的时候,这增强了该物种对春季霜冻的脆弱性。
科学研究中普遍采用的方法是量化成年成熟树木的表现,通常用于评估收获潜力。即使是对未经管理的天然林中气候影响的树木年代或树木年轮调查,也主要只分析了树冠占主导地位的树木的近期生活史。这种方法阻碍了对气候变化对森林生产力和功能的影响的全面了解。一些专注于高度生长的研究表明,在早期发育阶段,最近再生的树木比较老的树木生长得更快,这可能是气候变暖的有益影响的结果。然而,缺乏对树种在早期发育阶段二次生长的长期趋势的明确评估。
最近的文献证据显示,不同生命史阶段的环境反应存在显著差异,对比了上层和下层树木对环境变化的反应相似的普遍假设。幼树根系发育较差,碳水化合物和水分储量较低,对干旱胁迫更敏感。土壤和大气湿度水平以及泉水输入已被发现是树苗生长的重要驱动因素。森林冠层缓冲了气候的时间变化,在夏季的几个月里,相对于冠层以上的条件,保持了凉爽和湿润的林下条件。因此,与幼树、亚冠层树木(树苗)相比,成熟的上冠层树木可能因直接暴露在不利条件下和更高的用水需求而受到更大的负面影响。矛盾的发现表明,与上层树木相比,在温暖的条件下,下层树木的生长减少幅度更大,这表明气候与其他非生物和生物因素之间存在复杂的相互作用。
幼树和更普遍的林下树木在光和土壤资源方面面临着激烈的竞争,这可能会提高它们对温度和湿度限制的敏感性。森林结构和成分的异质性可能会缓冲或加剧树木对气候变化的敏感性。例如,与单物种林分相比,以高物种多样性为特征的森林群落可能较少受到干旱条件的影响。不同的物种占据着独特的生态位,因此减少了总体资源需求。因此,树苗的生长和存活率可以在d澳门手机赌博样化的环境中提高。然而,由于涉及许多相互作用的过程,在d澳门手机赌博样化的林分中并不总是观察到更高生产力的直观模式。鉴于幼树表现在塑造未管理林分的未来结构和生产力方面的相对重要性,我们认为我们对林下树木的生长动态和气候敏感性的理解需要进一步探索。
源自人类发展的污染物的沉积也对森林生长产生了强烈影响。例如,在中欧,20世纪中期燃煤工业的扩张导致森林地区硫和氮化合物的沉积急剧增加。随之而来的森林土壤酸化,以及对光合植物器官的损害,严重影响了树木的生理和生长。反过来,这些影响改变了树木对气候变化的敏感性。在某些情况下,随后森林结构和成分的变化导致人们错误地将最近的增长归因于气候变暖。
考虑到影响树木人口统计表现的众多结构力,需要一种多元创新的分析方法。我们提出了一个灵活的框架,以适应复杂的相互作用和非线性关系。我们使用了一个庞大的现有数据集,该数据集是从广泛的欧洲森林监测地块网络(www.remoteforests.org)中汇编而成的,该网络每年提供35000多棵树的年轮测量结果。挪威云杉和欧洲山毛榉这两个具有重要经济和生态意义的物种尤其有代表性。远程调查地块主要建立在未经管理的森林中。缺乏以往的管理有助于更直接地评估气候与气候的关系。我们调查了挪威云杉和欧洲山毛榉生长动态的驱动因素及其长期趋势,重点关注现已成熟的树木的早期生命阶段。我们假设:
(1) 由于气孔调节的种间差异,相对于山毛榉,云杉的生长对水分限制更敏感,但这种敏感性可以在d澳门手机赌博样化的林分中通过物种间资源的有效分配得到改善;
(2) 树苗的生长速度随着温度的升高而增加,由于时间变暖的趋势,导致物种生产力的长期提高。
04
—
结果
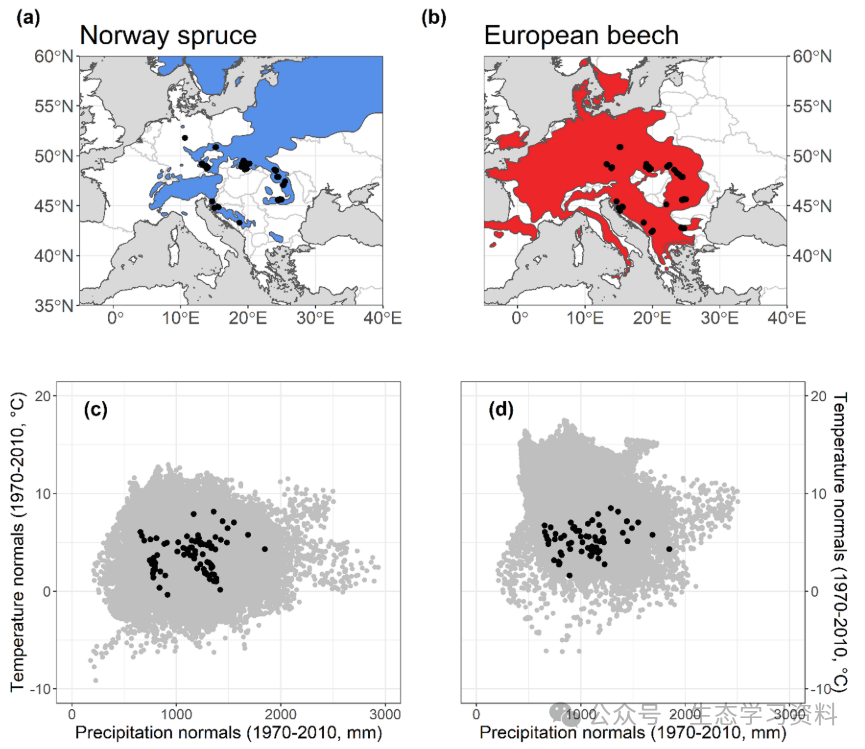
图1. REMOTE永久绘图网络。面板(a)和(b):显示欧洲森林论坛计划量化的云杉和山毛榉(阴影区域)当前地理分布的地图(Caudullo等人,2021)。黑色数据点对应于研究图的位置。面板(c)和(d):云杉和山毛榉的气候包络线。灰色数据点表示从网格气候数据集(CRU-TS 4.04)子集到每个物种的地理范围的所有像素(0.5度单元大小)的40年气候法线(如图a和b所示)。黑点是远程测量图的法线。降水正常值计算为1970-2010年期间年总降水量的40年平均值。温度法线定义为1970-2010年的年平均温度。请注意,在生长季节(4月至9月),云杉地的平均温度和气候水分平衡(CWB,降水量-潜在蒸散量)分别为12℃和220.90 mm,而山毛榉地的平均气温和气候水分均衡分别为13.58℃和161.57 mm。
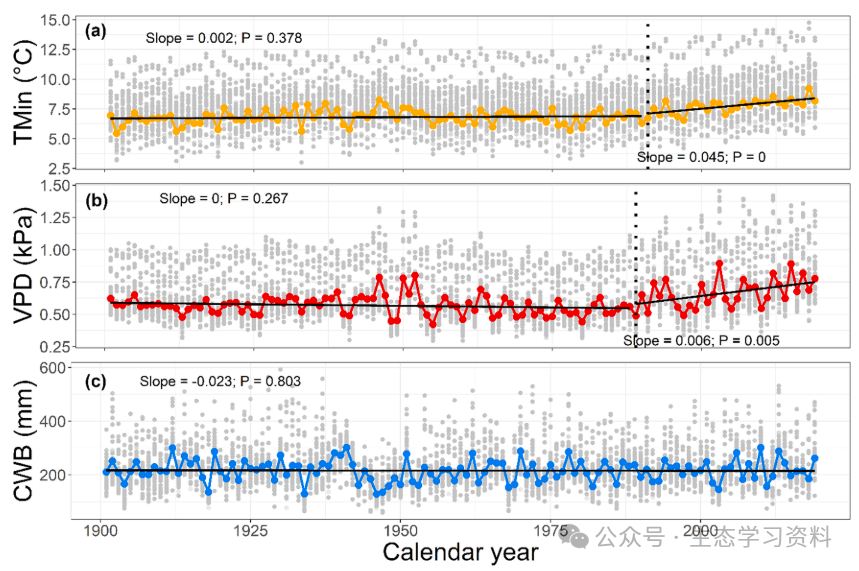
图2. 研究区域内的气候趋势。1901-2019年生长季节最低温度(TMin)、夏季蒸汽压亏缺(VPD)和气候水分平衡(CWB)的时间序列。TMin计算为时间序列中每年的生长季节(4月至9月)月平均最低温度。时间序列基于从CRU-TS 4.04数据集中提取的我们绘图位置的气候平均值。粗体线表示时间序列趋势显著变化前后的时间趋势(使用Pettitt检验检测)。断点由垂直虚线表示。就温度而言,1991年被确定为趋势从不显著变为积极的一年。对于蒸汽压不足,这种转变发生在1989年。没有检测到气候水分平衡的显著断点。灰点是绘图级别的值。还报告了针对时间相关数据校正的Sen斜率和p值。
表1. 1901-2010年期间特定物种GAMM预测BAI的输出。对于线性预测因子(拦截和释放),Estimate是估计的效应大小,Std.Error是标准误差。还显示T值和p值。对于光滑项,edf是估计的自由度,F是F统计量。显著性用p值表示,其中***表示p<=0.001,**p<=0.01,*p<=0.05,NS不显著。还显示了调整后的r平方值(Adj.R2)和观测次数(N)。
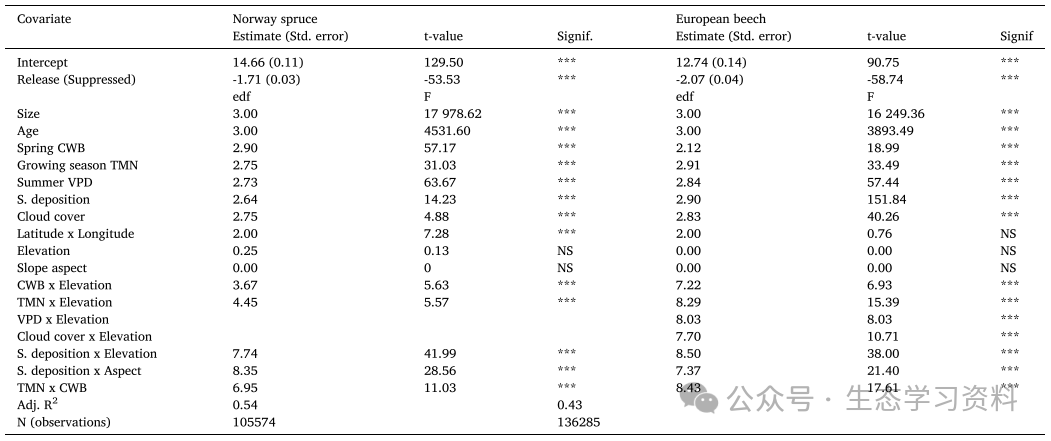
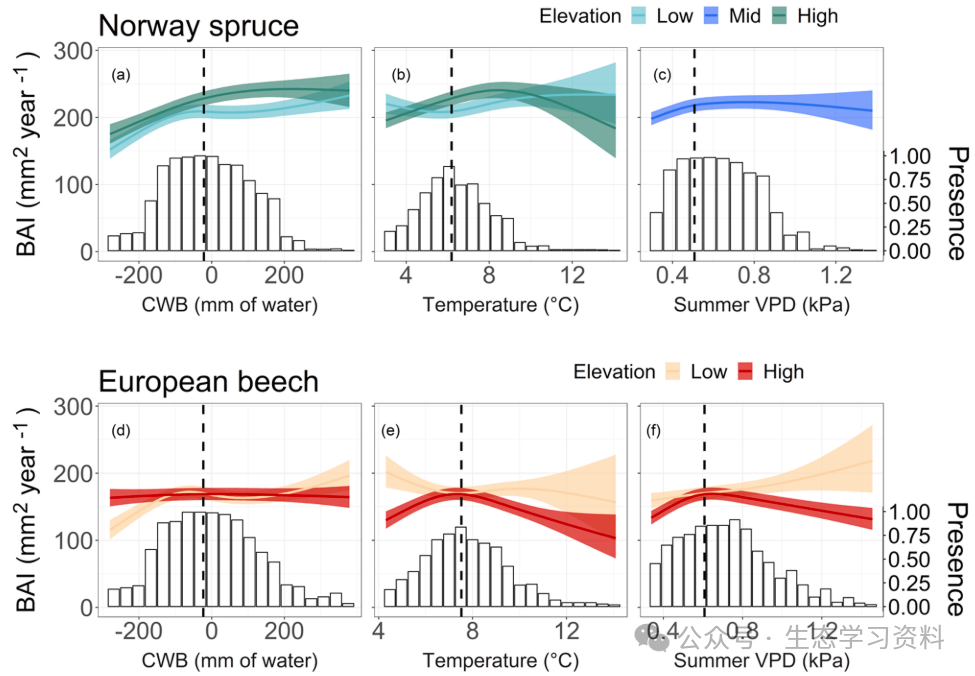
图3. 气候对两个重点物种在海拔梯度上幼年生长的影响。生长反应是由适合1901–2010年时期的物种特异性GAMM产生的。实线表示反向变换的预测基底面积增量(BAI)。阴影区域描绘了95%的置信区间。显示了两个海拔范围的预测BAI,分别对应于每个物种观测到的海拔的第10个百分位数(“低”,云杉和山毛榉分别为1235 m和732 m)和第90个百分位位数(“高”,云杉、山毛榉分别为1558 m和1258 m)。在非显著相互作用项的情况下(面板c),BAI是根据观测到的海拔中值(“Mid”,即云杉的1388m)进行预测的。模型中的其他解释因素(等式(5))保持在其相应的观测到的中间水平。虚线表示观测到的解释变量的中值。直方图(“存在”)表示物种分布数据,计算为气候区间给定物种存在的地块总数的分数。
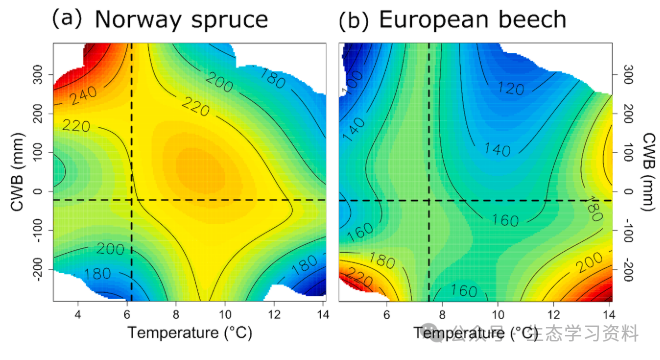
图4. 热图显示了云杉(a)和山毛榉(b)受最低温度和气候水分平衡(CWB)影响的基底面积增量(BAI)的变化。这些影响源自1901-2010年期间的GAMM(等式(5))。预测增长率显示为平滑表面,代表年度BAI的变化。BAI被缩放为从低(蓝色)到高(红色)的颜色梯度。等高线表示恒定BAI。其他模型因子保持在观察到的中值水平。请注意,就海拔而言,这对应于云杉的1388米和山毛榉的1061米。虚线表示观测到的最低温度或CWB的中值。
表2. 物种特异性GAMM预测1970-2010年BAI的输出。对于线性预测因子(拦截和释放),Estimate是估计的效应大小,Std.Error是标准误差。还显示T值和p值。对于光滑项,edf是估计的自由度,F是F统计量。显著性用p值表示,其中***表示p<=0.001,**p<=0.01,*p<=0.05,NS不显著。还显示了调整后的r平方值(Adj.R2)和观测次数(N)。
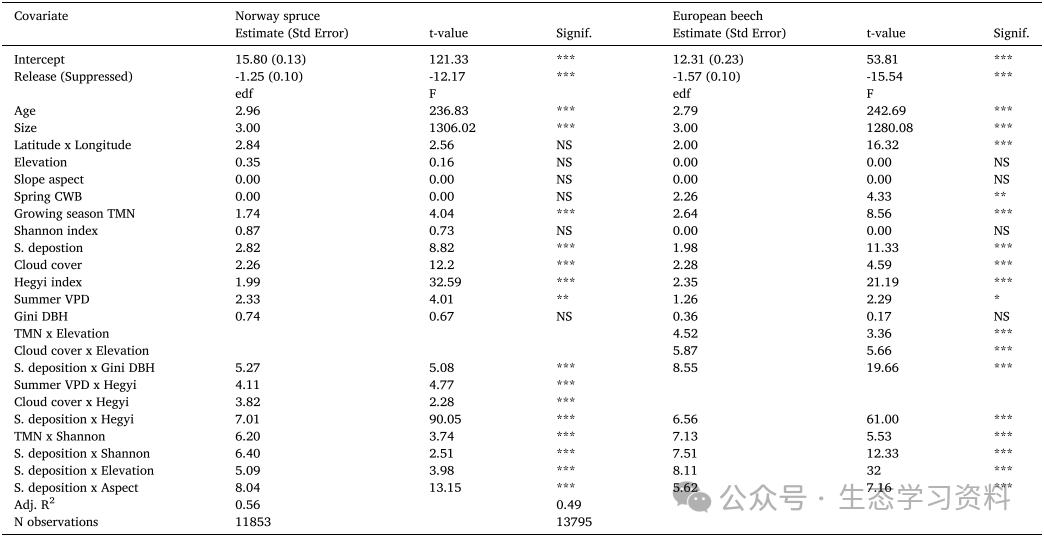

图5. 1970-2010年期间,平均生长季节最低温度和物种多样性之间的相互作用对特定物种GAMM幼树生长的影响(Eq.(6))。预测的生长速率显示为平滑表面,代表年基底面积增量(BAI)的变化。BAI被缩放为从低(蓝色)到高(红色)的颜色梯度。等高线表示恒定BAI。香农指数量化了单个目标树的调查邻域内的成分多样性。其他模型的因子保持在观察到的中值水平。虚线表示按物种划分的最低温度或香农指数值的中值。
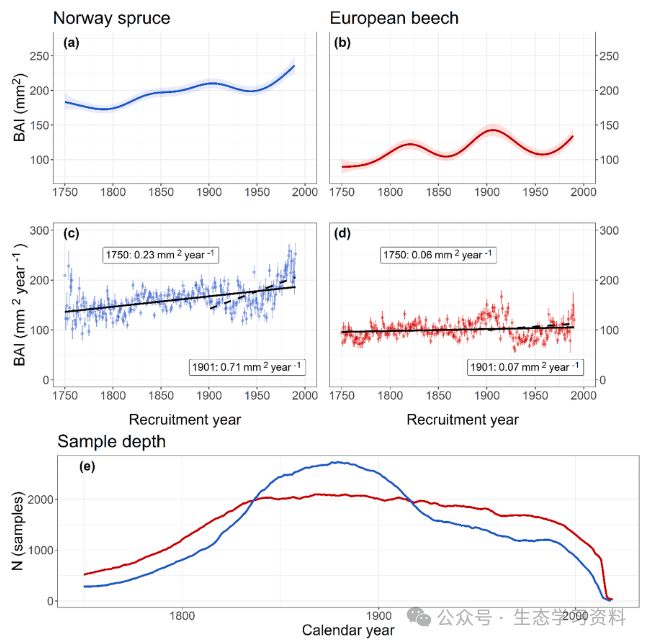
图6. 挪威云杉和欧洲山毛榉幼树生长的时间趋势。上图(a,b):GAMM的平均预测BAI,置信区间为95%(阴影区域)。中间面板(c,d):泰森斜率(趋势)估计。数据点显示了按招募年份和物种划分的BAI中值。误差条指定引导的95%置信区间。实线和虚线分别代表1750年和1901年之后招募的树木的BAI的时间趋势。除1901-2010年山毛榉外,所有趋势在α=0.05时都是显著的(Mann-Kendall趋势测试)。底部面板:样本深度,即按日历年划分的观测次数(树木年轮)。
05
—
结论
在这项研究中,我们将回归方程与个体树木的数据进行拟合,对比更传统的树木生态学方法,这些方法通常使用某种形式的标准化和聚合数据来调查种群水平对气候的反应。我们认为,个体树方法可以明确说明多种因素对生长的单独影响,包括个体发育和竞争,以及驱动因素之间的相互作用。我们的分析是现象学的,因此我们只能推测调节植物酚学的潜在生化过程的性质、形成层细胞的活性、生长的时间持续时间以及碳分配策略,这些策略可能会不同地促进木材的快速形成和耐应力。需要进一步的实验工作来量化这些关系和树木生长的基本限制。然而,我们的研究结果表明,变暖导致的潜在生长加速最终可能受到固有的基于生理的热阈值的限制,这与先前的工作一致。我们还认为,山毛榉和云杉在水分关系和耐受水分限制能力方面的差异很可能是由不同的生活史因素造成的。例如,有证据表明,与山毛榉相比,云杉以牺牲耐旱性为代价,优先考虑碳水化合物的生长。这些结果对森林通过温度介导的生物量增长和碳储存来缓解气候变化的能力具有影响。
阅读
839
| 我也说两句 |
| E-File帐号:用户名: 密码: [注册] |
| 评论:(内容不能超过500字。) |
*评论内容将在30分钟以后显示! |
| 版权声明: 1.依据《服务条款》,本网页发布的原创作品,版权归发布者(即注册用户)所有;本网页发布的转载作品,由发布者按照互联网精神进行分享,遵守相关法律法规,无商业获利行为,无版权纠纷。 2.本网页是第三方信息存储空间,阿酷公司是网络服务提供者,服务对象为注册用户。该项服务免费,阿酷公司不向注册用户收取任何费用。 名称:阿酷(北京)科技发展有限公司 联系人:李女士,QQ468780427 网络地址:www.arkoo.com 3.本网页参与各方的所有行为,完全遵守《信息网络传播权保护条例》。如有侵权行为,请权利人通知阿酷公司,阿酷公司将根据本条例第二十二条规定删除侵权作品。 |

协会微信公众号